
PENDAHULUAN
Seperti makhluk hidup melalui siklus hidup, setiap sel melalui siklus sel, tahap –tahap yang teratur selama pertumbuhan sel, persiapan untuk pembelahan, dan pembelahan menjadi dua sel anak, yang mengulang tahap-tahap tersebut. Eukariot sel tunggal yang hidup bebas, seperti Amoeba, siklus sel penting untuk keabadiannya. Dengan pembelahan sel tersebut, mereka menyebarkan informasi genetik kepada sel anaknya, yang sama seperti disebarkan pada jutaan generasinya. Dalam tumbuhan dan hewan multiseluler, sejumlah sel terusmenerus tumbuh dan membelah untuk kehidupan organisme tersebut. Pada tumbuhan terdapat pada sel-sel di ujung akar yang selalu memasuki kedalaman tanah. Pada hewan, sel-sel tersebut terdapat pada usus halus saluran pencernaan dan secara terus-menerus menghasilkan sel baru yang mengelupas bersamaan dengan makanan yang dicerna.
Pada sebagian besar organisme multiseluler, terjadi siklus sel sangat lambat atau berhenti membelah dan tetap, hal ini terjadi pada bagian tahap kehidupan seseorang yang berumur tua atau sakit atau memasuki kematian. Sel yang termasuk disini adalah sel otot, dan sel saraf pada hewan, sedangkan pada tumbuhan adalah sel-sel pembuluh yang mengatur pembentukan gula.
Pada dasarnya, pembelahan sel dibedakan menjadi 2 macam, yaitu pembelahan sel secara langsungdan secara tak langsung. Pembelahan sel secara langsung jika proses pembelahan tidak didahului dengan pembentukan gelendong pembelahan dan penampakan kromosom, disebutam itos is. Adapun pembelahan sel secara tak langsung jika proses pembelahan didahului dengan pembentukan gelondong pembelahan dan penampakan kromosom. Pembelahan secara tidak langsung meliputi pembelahan mitosis dan pembelahan meiosis.
Pembelahan amitosis terjadi pada bakteri, Protozoa, dan ganggang bersel satu. Proses pembelahan ini tidak melalui tahapan-tahapan pembelahan. Satu sel induk akan membelah secara langsung menjadi dua, dua menjadi empat, empat menjadi delapan, dan seterusnya hingga sel tersebut bertambah banyak. Proses pembelahan langsung didahului oleh pembelahan inti menjadi dua, diikuti oleh pembelahan sitoplasma dan akhirnya sel terbagi menjadi dua sel anak.
Tujuan:
1. Untuk mengetahui lebih jelas tahapan- tahapan pembelahan sel secara mitosis.
2. Untuk mengetahui lebih jelas tahapan- tahapan pembelahan sel secara meiosis.
3. Untuk mengetahui fungsi dari siklus sel.
BAB II
PEMBAHASAN
Siklus sel adalah rangkaian peristiwa perkembangan sel dengan urutan tertentu yang akan kembali pada tahap semula. Siklus sel terdiri dari dua tahap, yaitu interfase dan tahap mitotik.Interfase merupakan fase antara mitosis yang satu dengan mitosis berikutnya. Interfase terdiri atas tiga tahap yaitu GI (awal dari interfase), tahap S, dan tahap G2 (akhir dari interfase). Pada fase S terjadi sintesis atau duplikasi DNA.
Kromosom memiliki peranan yang sangat penting bagi keberlangsungan suatu makhluk hidup, karena kromosom merupakan alat pengangkutan gen – gen yang dipindahkan dari suatu sel ke sel yang lainnya, dan dari generasi yang satu ke generasi yang lainnya. Pemindahan gen – gen tersebut dilakukan pada saat siklus sel.
Siklus sel juga merupakan proses duplikasi secara akurat untuk menghasilkan jumlah DNA kromosom yang cukup banyak dan mendukung segregasi untuk menghasilkan dua sel anakan yang identik secara genetik atau empat sel anak yang hanya memiliki separuh jumlah kromosom dari sel sebelumnya. Proses ini berlangsung terus-menerus dan berulang - ulang (siklik).
Siklus sel dapat dibedakan menjadi empat fase yaitu gap – 1 (fase G1), sintesis (fase S), dan gap – 2 (G2). Dan mitosis atau meiosis (fase M).
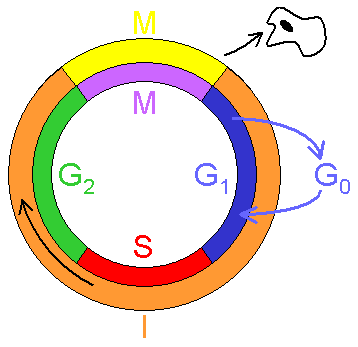
Fungsi utama dari siklus sel adalah menduplikat sejumlah DNA di dalam kromosom dengan tepat, kemudian
membelah menjadi dua sel anak yang identik. Proses ini merupakan dua fase utama dari siklus sel. Proses
duplikasi DNA terjadi pada fase S (S= sintesis), yang menghabiskan 10-12 jam dan merupakan separuh waktu
siklus sel pada tipe sel mamalia. Setelah fase S, terjadi pemisahan kromosom dan pembelahan sel pada fase M
(M=Mitotik), yang membutuhkan waktu lebih sedikit ( kurang dari satu jam pada sel mamalia). Mitosis terjadi
Pada fase M yang dimulai dengan kromosom yang terkondensasi. Fase mitotik merupakan tempat terjadinyi
mitosis. Kondensasi kromosom dan pembatasan kromosom replikan terjadi dalam mitosis(Alberts, 2002).
Fase mitosis pada umumnya merupakan fase siklus sel yang memiliki waktu yang paling pendek dibandingkan dengan fase G1, G2, sintesis, dan juga meiosis. Dalam siklus sel terjadi proses pembelahan sel yaitu pembelahan mitosis dan pembelahan meiosis. Perilaku kromosom berbeda pada saat mitosis dan pada saat meiosis. Pembelahan mitosis adalah pembelahan sel dimana terjadi pembelahan dan pembagian sel nukleus beserta kromosom – kromosom di dalamnya.
Nukleus yang tadinya hanya ada satu akan menjadi dua nukleus anakan yang sama. Proses pembelahan nukleus dinamakan kariokinesis. Kariokinesis ini akan diikuti oleh pembelahan sel, sehigga satu sel induk akan menjadi dua sel anakan. Proses pembelahan sel ini dinamakan sitokinesis. Kariokinesis dan sitokinesis yang terjadi secara berkesinambungan menyebabkaninformasi genetik di dalam semua sel somatis suatu individu selalu tetap.
Pembelahan meiosis merupakan pembelahan sel yang spesifik karena hanya terjadi pada saat pembentukan gamet. Jumlah kromosom yang diturunkan kepada keturunannya menjadi separuhnya saja haploid (n) dari yang tadinya diploid (2n) yang disebut juga sebagai pembelahan reduksi. Namun, setelah fertilisasi (bergabungnya antar gamet haploid) akan terbentuk kembali zigot yang diploid (2n). informasi genetik dari tetua yang satu akan memisah secara teratur ke dalam masing – masing gamet yang nantinya akan tercampur dengan informasi genetik dari gamet yang berasal dari tetua yang lainnya.
MITOSIS
Pembelahan mitosis adalah pembelahan sel dimana terjadi pembelahan dan pembagian sel nukleus beserta kromosom – kromosom di dalamnya. Nukleus yang tadinya hanya ada satu akan menjadi dua nukleus anakan yang sama. Proses pembelahan nukleus dinamakan kariokinesis. Kariokinesis ini akan diikuti oleh pembelahan sel, sehigga satu sel tetua akan menjadi dua sel anakan. Proses pembelahan sel ini dinamakan sitokinesis. Kariokinesis dan sitokinesis yang terjadi secara berkesinambungan menyebabkan informasi genetik di dalam semua sel somatis suatu individu selalu tetap.
Siklus mitotik sebuah sel dapat dibedakan menjadi dua bagian, yaitu stadium istirahat (interfase) dan stadium pembelahan (mitosis). Interfase sendiri dapat dibedakankembali menjadi tiga fase, yaitu fase gap satu (G1), fase sintesis (S), dan fase gap 2 (G2). Sedangkan stadium mitosis sendiri juga dibedakan kembali menjadi empat fase, yaitu profase, metafase, anafase, dan telofase. Namun ada juga yang menambahkan metakinesis diantara profase dan metafase.
Pembelahan mitosis menghasilkan sel anakan yang jumlah kromosomnya sama dengan jumlah kromosom sel induknya, pembelahan mitosis terjadi pada sel somatic (sel penyusun tubuh). Sel – sel tersebut juga memiliki kemampuan yang berbeda – beda dalam melakukan pembelahannya, ada sel – sel yang mampu melakukan pembelahan secara cepat, ada yang lambat dan ada juga yang tidak mengalami pembelahan sama sekalisetelah melewati masa pertumbuhan tertentu, misalnya sel – sel germinatikum kulit mampu melakukan pembelahan yang sangat cepat untuk menggantikan sel – sel kulit yang rusak atau mati. Akan tetapi sel – sel yang ada pada organ hati melakukan pembelahan dalam waktu tahunan, atau sel – sel saraf pada jaringan saraf yang sama sekali tidak tidak mampu melakukan pembelahan setelah usia tertentu. Sementara itu beberapa jenis bakteri mampu melakukan pembelahan hanya dalam hitungan jam, sehingga haya dalam waktu beberapa jam saja dapat dihasilkan ribuan, bahkan jutaan sel bakteri. Sama dnegan bakteri, protozoa bersel tunggal mampu melakukan pembelahan hanya dalam waktu singkat, misalkan amoeba, paramecium, didinium, dan euglena.
Pada jurnal yang berjudul “THE ROLE OF ADENOSINE 3',5'-CYCLIC MONOPHOSPHATE IN THE REGULATION OF MAMMALIAN CELL DIVISION” dalam jurnal ini dijelaskan bahwa Kelaparan dapat memegang proliferasi sel-sel di bagian yang berbeda dari siklus sel. Dalam rangka untuk menemukan di mana dalam siklus sel sel-sel ini diadakan. Peristiwa ini diukur sebagai fungsi waktu setelah memulai makan: (l) indeks mitosis, dan (2) indeks DNA sintetis (jumlah sel dalam DNA sintesis 1 jam setelah injeksi Ha-timidin). Durasi periode DNA sintetis sel (S) diukur, memungkinkan deskripsi yang lebih tepat dari siklus sel. Analisis epitel duodenum dan esofagus menunjukkan makan yang memulai pembelahan sel oleh sel-sel merangsang dari bagian G1 dari siklus mitosis dalam duodenum. Dalam kerongkongan beberapa sel-sel baik berhenti atau melambat di G 1, dan kelompok sel di G2. Makan secara simultan merangsang kelompok sel keduanya; bergerak mantan ke S, yang terakhir ke dalam mitosis. Periode S pada hewan kelaparan adalah sedikit lebih lama dari itu pada hewan biasanya makan tapi ekstensi dapat dikaitkan dengan suhu tubuh sedikit menurun.
Pada jurnal yang berjudul “TELAAH BEBERAPA FUNGSI TITIK-UJI SIKLUS PEMBELAHAN SEL FASE G1 DAN S DARI INHIBITOR KINASE- BERGANTUNG SIKLIN SICI” Untuk dapat menentukan proses yang dipantau oleh fungsi titik-uji dari Sic1, penelitian ini telah dilakukan telaah terhadap pengaruh pelesapan gen SIC1 terhadap beberapa proses fase G1dan S yang telah diketahui. Telaah ini dimulai dengan meneliti respon sel Δsic1 terhadap kerusakan DNA akibat sinar UV dan sinar-γ. Pada sel mamalia, sebagai respon terhadap kerusakan DNA akibat penyinaran-γ, protein penghambat tumor p53 akan menginduksi transkripsi suatu CKI (=Cyclin dependent kinase inhibitor = inhibitor kinase-bergantung-siklin) mamalia, yakni p21CIP1. p21CIP1 yang terinduksi sintesisnya, akan menghentikan siklus sel, sehingga memberi waktu bagi sel untuk memperbaiki kerusakan pada DNA, atau bila kerusakan tak dapat diperbaiki, menyebabkan sel mengalami apoptosis.
Pada sel – sel organisme multiseluler, proses pembelahan sel memiliki tahap – tahap tertentu yang disebut siklus sel. Sel – sel tubuh yang aktif melakukan pembelahan memiliki siklus sel yang lengkap. Siklus sel tersebut dibedakan menjadi dua fase(tahap ) utama, yaitu interfase dan mitosis. Interfase terdiri atas 3 fase yaitu fase G, ( growth atau gap), fase S (synthesis), fase G2(growth atau Gap2).
Pembelahan mitosis dibedakan atas dua fase, yaitu kariokinesis dan sitokinesis, kariokinesis adalah proses pembagian materi inti yang terdiri dari beberapa fase, yaitu Profase, Metafase, dan Telofase. Sedangkan sitokinesis adalah proses pembagian sitoplasma kepada dua sel anak hasil pembelahan.
1. Kariokinesis
Kariokinesis selama mitosis menunjukkan cirri yang berbeda – beda pada tiap fasenya. Beberapa aspek yang dapat dipelajari selama proses pembagian materi inti berlangsung adalah berubah – ubah pada struktur kromosom,membran inti, mikro tubulus dan sentriol. Cirri dari tiap fase pada kariokinesis adalah:
a) Profase
1. Benang – benang kromatin berubah menjadi kromosom. Kemudian setiap kromosom membelah menjadi kromatid dengan satu sentromer.
2. Dinding inti (nucleus) dan anak inti (nucleolus) menghilang.
3. Pasangan sentriol yang terdapat dalam sentrosom berpisah dan bergerak menuju kutub yang
berlawanan.
4. Serat – serat gelendong atau benang – benang spindle terbentuk diantara kedua kutub
pembelahan.
b) Metafase
Setiap kromosom yang terdiri dari sepasang kromatida menuju ketengah sel dan berkumpul
pada bidang pembelahan (bidang ekuator), dan menggantung pada serat gelendong melalui
sentromer atau kinetokor.
c)Anaphase
Sentromer dari setiap kromosom membelah menjadi dua dengan masing – masing satu kromatida.
Kemudian setiap kromatida berpisah dengan pasangannya dan menuju kekutub yang berlawanan.
Pada akhir nanfase, semua kroatida sampai pada kutub masing – masing.
d)Telofase
Pada telofase terjadi peristiwa berikut:
1. Kromatida yang berada jpada kutub berubah menjasadi benang-benangkromatin kembali.
2. Terbentuk kembali dinding inti dan nucleolus membentuk dua inti baru.
3. Serat – serat gelendong menghilang.
4. Terjadi pembelahan sitoplasma (sitokenesis) menjadi dua bagian, dan terbentuk membrane sel pemisah ditengah bidang pembelahan. Akhirnya , terbentuk dua sel anak yang mempunyai jumlah kromosom yang sama dengan kromosom induknya.
2 Sitokinesis
Selama sitokinesis berlangsung, sitoplasma sel hewan dibagi menjadi dua melalui terbentuknya cincin kontraktil yang terbentuk oleh aktin dan miosin pada bagian tengah sel. Cincin kontraktil ini menyebabkan terbentuknya alur pembelahan yang akhirnya akan menghasilkan dua sel anak. Masing – masing sel anak yang terbentuk ini mengandung inti sel, beserta organel – organel selnya. Pada tumbuhan, sitokinesis ditandai dengan terbentuknya dinding pemisah ditengah – tengah sel. Tahap sitokinesis ini biasanya dimasukkan dalam tahap telofase.
Pada jurnal yang berjudul “Delay of HeLa Cell Cleavage into Interphase Using Dihydrocytochalasin B: Retention of a Postmitotic Spindle and Telophase Disc Correlates with Synchronous Cleavage Recovery ”.Di dalam jurnal ini dijelaskan tentang molekul sinyal yang menentukan posisi dan waktu alur pembelahan selama mamalia mengalami sitokenesis sel. Kami
telah mempelajari secara rinci efek dihydrocytochalasin B (DCB), obat yang mengganggu aktin perakitan, pada spesifik peristiwa mitosis akhir dalam sel HeLa sinkron.
Di dalam jurnal ini juga menyimpulkan bahwa pembelahan sel dapat terjadi di G1 dan karena itu peristiwa metabolik independen dari mitosis. Para telofase dipertahankan
disk mungkin memang sinyal posisi alur pembentukan, pembelahan G1 seperti terjadi hanya dalam posisi dimana disk dipertahankan mendasari korteks sel. Para protokol kami menjelaskan sekarang harus memungkinkan pengembangan dari sebuah sistem model untuk studi sel mamalia pembelahan sebagai peristiwa sinkron independen dari mitosis.
MEIOSIS
Pada tahun 1883 atau empat tahun setelah mitosis dapat dijelaskan, Edouard van Beneden menemukan bahwa telur cacing Ascaris mengandung kromosom hanya separuh jumlah kromosom yang terdapat di dalam sel-sel somatisnya. Ia kemudian dengan tepat dapat menginterpretasikan hal itu sebagai akibat terjadinya suatu tipe pembelahan sel yang lain, yang disebut meiosis (meioun = pengurangan).
Meskipun demikian, Beneden salah menyimpulkan bahwa pada pembelahan meiosis seluruh kromosom paternal (kromosom dari tetua jantan) akan bergerak ke satu kutub sel dan seluruh kromosom maternal (kromosom dari tetua betina) bergerak ke kutub sel yang lain. Peristiwa yang benar adalah terjadi percampuran kromosom paternal dan maternal membentuk pasangan-pasangan kromosom homolog, yang kemudian disebarkan secara acak ke dalam sel-sel hasil meiosis.
Bila dibandingkan dengan mitosis, meiosis membutuhkan waktu yang jauh lebih panjang dengan proses yang lebih rumit. Meiosis dapat dibagi menjadi dua pembelahan nukleus (kariokinesis), yaitu meiosis I dan meiosis II. Pada meiosis I terjadi pengurangan jumlah kromosom menjadi setengah dari semula sehingga pembelahan ini sering juga disebut pembelahan reduksi. Jika sel yang mengalami meiosis adalah sebuah sel diploid, maka pada akhir meiosis II akan didapatkan empat buah sel yang masing-masing haploid. Hal ini karena kromosom hanya mengalami satu kali penggandaan, tetapi kariokinesisnya terjadi dua kali.
Tahap-tahap meiosis
Oleh karena meiosis dapat dibagi menjadi meiosis I dan meiosis II, maka tahaptahapnya terdiri atas profase I, metafase I, anafase I, telofase I, profase II, metafase II, anafase II, dan telofase II. Tahap-tahap meiosis II (profase II hingga telofase II) sebenarnya menyerupai tahap-tahap pada mitosis.
Profase I
Di antara tahap-tahap meiosis, profase I membutuhkan waktu paling panjang sehingga dapat dibagi lagi menjadi beberapa tahap, yaitu leptonema, zigonema, pakinema, diplonema, dan diakinesis.
Ø Leptonema (leptoten)
Seperti halnya pada profase awal mitosis, pada tahap meiosis yang paling awal ini setiap kromosom telah mengalami penggandaan menjadi kromatid kembar. Namun, kenampakan kromosom jika dilihat menggunakan mikroskop cahaya masih seperti benang tunggal yang tipis memanjang. Di sepanjang kromosom dijumpai sejumlah kromomir, berupa butiran-butiran padat dengan interval yang tidak beraturan.
Ø Zigonema (zigoten)
Tiap kromosom homolog (kromosom paternal dan maternal) berpasang-pasangan membentuk struktur bivalen. Proses berpasangannya sendiri dinamakan sinapsis. Oleh karena tiap kromosom telah mengalami penggandaan menjadi dua kromatid kembar,maka pada tiap bivalen terdapat empat kromatid kembar. Kompleks empat kromatid ini disebut tetrad.
Ø Pakinema (pakiten)
Pada pakinema kromosom untuk pertama kalinya dapat dilihat sebagai struktur yang telah mengalami penggandaan (bivalen atau tetrad). Peristiwa penting lainnya pada tahap ini adalah terjadinya pindah silang (crossing over), yaitu pertukaran materi genetik antara kromatid paternal dan kromatid maternal pasangannya.
Ø Diplonema (diploten)
Secara visual tempat terjadinya pindah silang dapat dilihat sebagai struktur yang dinamakan kiasma (jamak = kiasmata). Kecuali pada daerah-daerah kiasma ini, pasangan-pasangan kromatid nampak mulai saling memisah.
Ø Diakinesis
Kiasma bergeser ke ujung kromosom sehingga tempat ini sekarang tidak harus merupakan tempat terjadinya pindah silang. Tiap kromatid anggota tetrad makin memendek, menebal, dan bergerak ke arah bidang tengah sel. Nukleolus dan dinding nukleus menghilang. Mikrotubul / benang spindel yang keluar dari sentriol nampak kian memanjang dan akhirnya melekat pada kinetokor.
Metafase I
Struktur tetrad nampak makin jelas di bidang tengah sel. Di sinilah konfigurasi kromosom meiosis paling mudah dibedakan dengan kromosom metafase mitosis. Pada metafase mitosis tidak dijumnpai adanya struktur tetrad, tetapi hanya ada biad yang terdiri atas dua kromatid kembar.
Anafase I
Anggota tiap pasangan kromosom homolog (yang masing-masing terdiri atas dua kromatid kembar) bergerak ke arah kutub sel yang berlawanan. Dalam hal ini sentromir belum membelah sehingga kedua kromatid kembar masih terikat satu sama lain.
Telofase I
Anggota tiap pasangan kromosom homolog telah mencapai kutub sel yang berlawanan. Dinding nukleus mulai terbentuk kembali. Kadang-kadang telofase I diikuti oleh sitokinesis dan interfase singkat (tanpa penggandaan kromosom), tetapi seringkali langsung diteruskan ke meiosis II.
Meiosis II
Di atas telah dikatakan bahwa tahap-tahap meiosis II, mulai dari profase II hingga telofase II, menyerupai tahap-tahap pada mitosis. Namun, pada meiosis II hanya ada satu dari masing-masing pasangan kromosom homolog di dalam setiap nukleus. Jadi, di dalam tiap nukleus hanya ada kromosom paternal saja atau kromosom maternal saja untuk tiap nomor kromosom. Sebagai contoh, di dalam satu nukleus mungkin terdapat kromosom paternal untuk kromosom nomor 1, kromosom maternal untuk kromosom nomor 2, kromosom maternal untuk kromosom nomor 3, dan seterusnya. Nukleus lainnya akan membawa kombinasi kromosom yang lain pula.
Telofase II akan diikuti oleh sitokinesis yang menghasilkan empat sel haploid. Di dalam nukleus masing-masing sel ini terdapat satu anggota untuk setiap pasangan kromosom homolog. Jadi, kalau pada telofase I (dan sebelumnya, anafase I) terjadi pemisahan kromosom homolog, pada telofase II (dan anafase II) terjadi pemisahan kromatid.
Gametogenesis pada hewan
Dengan berakhirnya meiosis tidak serta-merta dapat dikatakan bahwa gamet telah terbentuk. Meiosis hanya menghasilkan empat buah sel yang masing-masing haploid. nSel-sel ini masih memerlukan proses pematangan untuk dapat berkembang menjdi gamet. Pembelahan meiosis yang diikuti oleh pematangan sel-sel haploid menjadi gamet fungsional dinamakan gametogenesis.
Pada hewan yang berkembang biak secara seksual dapat dibedakan antara gametogenesis pada individu jantan dan gametogenesis pada individu betina. Gamet pada individu jantan disebut spermatozoon (jamak = spermatozoa) sehingga proses pembentukannya dinamakan spermatogenesis. Demikian pula, karena gamet betina disebut ovum (jamak = ova), maka gametogenesis pada jenis kelamin ini dinamakan oogenesis.
Spermatogenesis
Spermatogenesis dimulai pada saat individu yang bersangkutan mencapai matang kelamin (pubertas). Prosesnya berlangsung di dalam testes, tepatnya di dalam suatu tabung melengkung yang disebut tubulus seminiferus. Di sekeliling tabung ini terdapat spermatogonium (jamak = spermatogonia), yaitu sel-sel somatis khusus yang nantinya akan mengalami meiosis untuk menghasilkan spermatozoa.
Pada awalnya spermatogonium (diploid) memperbanyak diri melalui pembelahan mitosis berkali-kali. Pada waktu tertentu mitosis akan terhenti; spermatogonium membesar dan berdiferensiasi menjadi spermatosit primer, yang masih diploid juga.
Spermatosit primer kemudian mengalami meiosis I untuk menghasilkan spermatosit sekunder, yang dilanjutkan dengan meiosis II untuk menghasilkan empat buah spermatid yang masing-masing haploid. Akhirnya, spermatid berdiferensiasi menjadi spermatozoon yang matang.
Oogenesis
Bila dibandingkan dengan spermatogenesis, oogenesis relatif agak lebih rumit. Proses ini dimulai sejak awal tahap perkembangan embrio ketika sekelompok sel yang disebut galur sel germinal (germ cell line) memasuki ovarium yang sedang berkembang. Galur sel ini kemudian berkembang menjadi sel-sel somatis khusus yang disebut oogonium (jamak = oogonia).
Oogonium (diploid) memperbanyak diri dengan sangat cepat melalui pembelahan mitosis berkali-kali, dan akhirnya berdiferensiasi menjadi oosit primer, yang masih diploid juga. Oosit primer kemudian mengalami meiosis I tetapi tertahan pada tahap diplonema hingga saat matang kelamin. Selama kurun waktu ini oosit primer mengalami berbagai perubahan sehubungan dengan persiapan penyelesaian meiosis dan fertilisasi, serta mengumpulkan sejumlah besar bahan makanan untuk perkembangan awal embrio. Untuk melindungi diri dari kerusakan mekanis, oosit primer diselubungi oleh selaput yang dinamakan folikel Graaf. Di bawah selaput ini terdapat granula kortikal yang membatasi pembuahan hanya oleh satu spermatozoon.
Oosit primer yang berhasil menyelesaikan meiosis I akan menghasilkan dua buah sel haploid, yang masing-masing mengandung satu anggota pasangan kromosom homolog dalam keadaan mengganda. Namun, sitokinesis tidak berlangsung simetris sehingga kedua sel tersebut sangat berbeda kandungan sitoplasmanya. Sel yang mendapatkan hampir seluruh sitoplasma dinamakan oosit sekunder, sedangkan sel satunya yang hanya mendapatkan sangat sedikit sitoplasma dinamakan badan polar.
Oosit sekunder keluar dari folikel Graaf untuk memasuki saluran telur (pada manusia: tuba falopi ; pada hewan: oviduktus). Proses pelepasan oosit sekunder dari folikel Graaf dinamakan ovulasi. Baik oosit sekunder maupun badan polar akan melanjutkan oogenesis ke tahap meiosis II. Lagi-lagi, oosit sekunder mengalami sitokinesis yang tidak simetris sehingga diperoleh satu sel yang besar (ovum) dan satu sel yang kecil (badan polar). Dengan demikian, pada akhir meiosis II dari sebuah oogonium akan diperoleh empat buah sel haploid, yang terdiri atas sebuah ovum (sel telur) dan tiga badan polar. Ketiga badan polar segera mengalami degenerasi karena hanya mengandung sedikit sekali sitoplasma dan organel yang diperlukan untuk melangsungkan metabolisme.
Meiosis II hanya akan selesai jika terjadi fertilisasi. Ovum yang tidak dibuahi akan mengalami degenerasi. Sebaliknya, jika ovum bertemu dengan spermatozoon akan terjadi penggabungan dua nukleus haploid sehingga terbentuk zigot diploid, yang kemudian turun dari tuba falopi / oviduktus menuju ke uterus.
Pada jurnal yang berjudul “Maturation Rate of Ovine Oocytes from Different Reproductive Status and Maturation Medium ” Penelitian ini bertujuan untuk menentukan jumlah folikel, kualitas oosit dan tingkat pematangan oosit dari pasangan ovarium dengan status reproduksi berbeda dalam dua media pematangan TCM- 199 sebagai kontrol dan CR1 aa sebagai pengobatan. Pemasangan ovarium diklasifikasikan menjadi empat kelompok: 1. Dengan korpus luteum indung telur (CL) dan folikel dominan (DF), 2. Ovarium dengan CL tanpa DF, 3. Ovarium dengan DF, tanpa CL, 4. Ovarium tanpa kedua CL dan DF. Hasil penelitian ini mengungkapkan bahwa jumlah terbesar dari folikel diamati dari ovarium dengan CL tanpa DF dan yang terendah dengan oosit dari ovarium dengan DF tanpa CL dalam TCM-199 dan CR1 aa.
Dalam rangkaian proses produksi embrio (pematangan/ maturasi, fertilisasi, dan kultur embrio) secara in vitro, dibutuhkan suatu lingkungan mikro yang dapat mendukung pertumbuhan dan perkembangan embrio. Salah satu faktor penting yang mempengaruhi perkembangan embrio secara in vitro adalah pemilihan medium yang tepat. Secara umum, medium untuk produksi embrio in vitro dibedakan menjadi medium kompleks dan sederhana (Gordon 1994). Tissue culture medium (TCM)-199 merupakan medium kompleks yang bersifat komersial dan telah digunakan untuk produksi embrio sapi (Sumantri et al. 1998; Boediono et al. 2003), domba (Gomez etal. 1998; Jaswandi et al. 2001) dan babi (Wang et al. 1997).
Pada jurnal yang berjudul “A mad partner for Shugoshin in meiosis ” Para cohesin centromeric Pelindung Shugoshin (SGO) dan spindle yang Mad2 pemeriksaan protein dikenal untuk bertindak di jalur yang berbeda untuk memastikan kromosom akurat segregasi selama pembelahan sel eukariotik. Dalam jurnal ini menjelaskan tak terfduga fisik interaksi antara vertebrata SGO dan Mad2, serta indikasi potensi mereka saling regulasi dalam meiosis. Kanker dan sindrom Down adalah dua hasil dari kromosom missegregation dalam mitosis dan meiosis, masing-masing. Selama siklus sel somatik, segregasi akurat tergantung tentang regulasi yang tepat dari kompleks cohesin yang terus direplikasi. Pengikatan Mad2 endogen untuk Xenopus SGO (satu-satunya Shugoshin protein dalam organisme ini) pada oosit katak dikonfirmasi bahwa interaksi ini juga terjadi pada sel meiosis.
BAB III
KESIMPULAN
Ø Fungsi utama dari siklus sel adalah menduplikat sejumlah DNA di dalam kromosom dengan tepat, kemudian membelah menjadi dua sel anak yang identik. Proses ini merupakan dua fase utama dari siklus sel.
Ø Mitosis merupakan pembelahan inti yang berhubungan dengan pembelahan sel somatik, dimana terdapat beberapa tahap didalamnya, yaitu: interfase, profase, metafase, anafase dan telofase.
Ø Meiosis dibagi menjadi meiosis I dan meiosis II, maka tahapannya terdiri dari atas profase I, metafase I, anafase I, telofase I, profase II, metafase II, anafase II, dan telofase II. Tahap- tahap meiosis II (profase II hingga telofase II) sebenarnya menyerupai tahapan pada mitosis.
DAFTAR PUSTAKA
Anonim. 2008. Cell and Cyle Cell. http://www.tutornext.com/short-answer-typequestions-
cell-cycle/9925. 7 November 2010.
Boediono, A, dkk. 2006. Maturation Rate of Ovine Oocytes from Different Reproductive Status and Maturation Medium. Vol 13 No 4.
Lima – Brito, J., H. Guedes – Pinto, G. E. Harrison, and J. S. Haeslop – Harrison. 1996.Chromosome identification and nuclear architecture in triticale x tritordeum F1 hybrids. Journal of Experimental Botany. 47 (297) : 583 – 588.
Amstrong, S. J. and Gareth H. J. 2003. Meiotic sytology and chromosome behaviour in wild – type arabidopsis thaliana. Journal of Experimental. 54 (380) : 1 – 10
Hongtao Yu.2011. A mad partner for Shugoshin in Meiosis. The EMBO Journal (2011) 30, 2759–2761. doi:10.1038/emboj.2011.217. Department of Pharmacology, Howard Hughes Medical Institute, University of Texas Southwestern Medical Center, Dallas, TX, USA.
Martineau, S.N,dkk. 1995. Delay of HeLa Cell Cleavage into Interphase Using Dihydrocytochalasin B: Retention of a Postmitotic Spindle and Telophase Disc Correlates with Synchronous Cleavage Recovery. Institut de Biologie Structurale-Jean-Pierre Ebel, 38027 Grenoble, France.
Monahan.M.T, and Abell C.W. 1973. The Role of Adenosine 3',5'-CyclicMonophosphat
In The Regulation Of Mamalian Cell Division. the Department of Human Biological Chemistry and Genetics, Division of Biochemistry, The University of Texas Medical Branch, Galveston, Texas.
Nugroho T Titania, 1999. Telaah Beberapa Fungsi Titik Uji Siklus Pembelahan Sel Fase G1 Dan S dari Inhibitor. Kinase Bergantung Siklin SIC. Jurnal Natur Indonesia. Vol 1191):1-11
Tidak ada komentar:
Posting Komentar